Authors: Sunita Chaudhary*, Jyoti Chauhan , Seema and Rekha Sodani
Abstract: Wounded plants accumulate numerous plant defensive compounds. Now-a-days, it is well accepted that plant secondary metabolites viz phenolic compound Terpenes and Alkaloids are involved in this plant defense system. The process of inducing resistance using elicitors is environmental friendly and is advantageous over the chemical based pesticides. It is like stimulation of the plant’s own defence system rather than on suppression of pathogens. The resistance developed in this way has long lasting effect. Secondary metabolites often play an important role in plant defense against herbivory and other interspecies defenses. This could be an alternative solution to reduce the use of pesticides and protect plant from herbivorous and act as anti feedant.
Introduction: Secondary metabolites are organic compounds that are not directly involved in the primary growth, development. Unlike primary metabolites, absence of secondary metabolites does not result in immediate death, but rather in long-term impairment of the organism's survivability, fecundity, or aesthetics, or perhaps in no significant change at all. Secondary metabolites are often restricted to a narrow set of species within a phylogenetic group. One important defining quality of secondary metabolites is their specificity. Usually, secondary metabolites are specific to an individual species. Research also shows that secondary metabolic can affect different species in varying ways. In the same forest, four separate species of arboreal marsupial folivores reacted differently to a secondary metabolite in eucalypts. This shows that differing types of secondary metabolites can be the split between two herbivore ecological niches. Additionally, certain species evolve to resist plant secondary metabolites and even use them for their own benefit. For example, monarch butterflies have evolved to be able to eat milkweed (Asclepias) despite the toxic secondary metabolite it contains. This ability additionally allows the butterfly and caterpillar to be toxic to other predators due to the high concentration of secondary metabolites consumed.
Many secondary metabolites found in plants have a role in defense against herbivores, pests and pathogens. In this article, a few examples are described and discussed, and some of the problems in determining the precise role(s) of such metabolites highlighted. The role of secondary metabolites in defense may involve deterrence andanti-feedant activity, toxicity or acting as precursors to physical defense systems. Many specialist herbivores and pathogens do not merely circumvent the deterrent or toxic effects of secondary metabolites but actually utilize these compounds as either host recognition cues or nutrients (or both). Their biochemistry is compared and contrasted. An enormous variety of secondary metabolites are derived from shikimic acid or aromatic amino acids, many of which have important roles in defense mechanisms. Several classes of secondary products are ‘induced’ by infection, wounding or herbivory, and examples of these are given. Genetic variation in the speed and extent of such induction may account, at least in part, for the difference between resistant and susceptible varieties. Both salicylates and jasmonates have been implicated as signals in such responses and in many other physiological processes, though their precise roles and interactions in signaling and development are not fully understood. The classic view that secondary constituents are waste products in plants have been replaced by one in which their value is assumed to be as a protection against herbivorous and microbial attack. Large increases in the amount of secondary metabolites can occur in stressed plants and those subject to mechanical damage or that caused by insects. In order to establish a protective role for a given metabolite, it is necessary to monitor concentrations over the life cycle of the plant, to survey plant populations, to determine specific localization within tissues and to carry out bioassays against insects and microorganisms. Synergy between toxins of the same class or of different classes is likely. Changes in secondary chemistry may also occur during ontogeny, and protection may be restricted to the most vulnerable plant organs. Finally, toxins may vary indiscriminately in their distribution within the plant or within populations and still provide protection.
Induced plant defense against insect herbivorous: Plant develop wide variety of defense strategies against herbivory
1. Constitutive defense response : These defense mechanism is always present. they are often species specific and exit in as stored compounds to reduce toxicity or precursors of active compound, that can easily activate upon damage.
2. Induced defense response: Initiated only after damage occurs. It require smaller investment of plant resources but must activate quickly to be effective.
Biosynthesis of secondary metabolites:
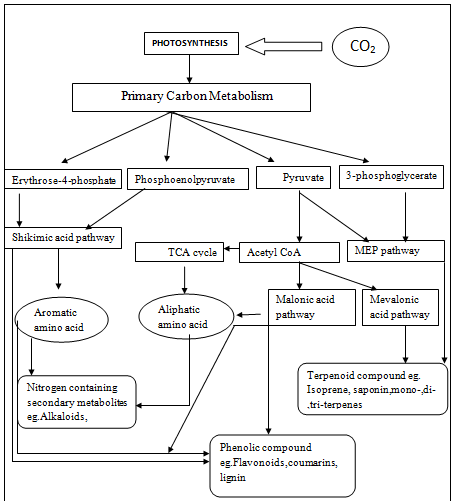
Conclusion: Secondary metabolites play significant role in plant defense against insect and, herbivorous. The apparent lack of primary function in the plant, combined with the observation that many secondary metabolites have specific negative impacts on other organisms such as herbivores and pathogens, leads to the hypothesis that they have evolved because of their protective value Production increases when a plant is attacked by herbivores or pathogens.
References:
Anonymous, 1985. Diseases of break crops. In: Rothamsted experimental station report for 1984. Harpenden : Lawes Agricultural Trust, 124 –125.
Atkin JC. 1979. Varietal susceptibility of potatoes to slug attack. Ph.D. thesis , University of Newcastle upon Tyne, UK.
Baayen RP, Sparnaaij J, Jansen J, Niemann GJ. 1991. Inheritance of resistance in carnation against Fusarium oxysporum f. sp. dianthi races 1 and 2, in relation to resistance components. Netherlands Journal of Plant Pathology 97: 73 –86.
Cheeke PR. 1989. Toxicants of plant origin , volume IV, phenolics. Boca Raton : CRC Press.
Chew FS. 1988. Biological effects of glucosinolates. In: CutlerHG, ed. Biologically active natural products. Washington DC : American Chemical Society, 155 –181.
Collinge DB, Slusarenko AJ. 1987. Plant gene expression in response to pathogens. Plant Molecular Biology 9: 389–410 .
Darvill AG, Albersheim P. 1984. Phytoalexins and their elicitors: a defence against microbial infection in plants. Annual Review of Plant Physiology 35 : 243–276.
Harborne JB. 1991. Role of secondary metabolites in chemical defence mechanisms in plants. In: ChadwickDJ, MarshJ, eds. Bioactive compounds from plants . Chichester : John Wiley and Sons, 126– 139.
Lieberei R, Biehl B, Giesemann A, Junqueira NTV. 1989. Cyanogenesis inhibits active defense reactions in plants. Plant Physiology 90 : 33–36.
About Author / Additional Info:
I am persuing ph.D degree from BHU varansi with Srf fellowship